Redigert av Dr. Giovanni Chetta
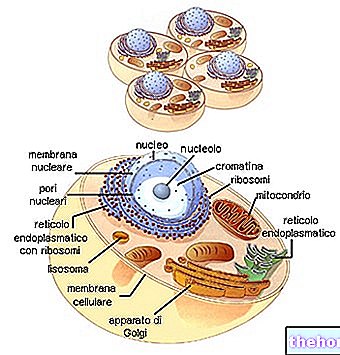
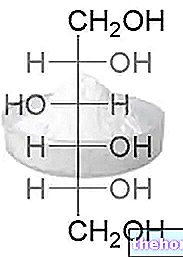
Det ekstracellulære nettverket
ECM kan derfor betraktes som et veldig komplekst nettverk der proteiner, PGS og GAG gir utallige funksjoner, inkludert strukturell støtte og regulering av hvert vev og organisk aktivitet. Global mobil homeostase bør betraktes som et kompleks av mekanismer som kan oppstå og utvikle seg inne i cellen eller utenfor ECM; i sistnevnte tilfelle kan cellen representere det mellomliggende eller siste målet. De ekstracellulære komponentene, i tillegg til å representere fysiske støttestrukturer for det cellulære stillaset, fungerer også som virkelige steder for initiering, utvikling og avslutning av vitale prosesser som angår både det endocellulære miljøet og organer og systemer. Vi står overfor et uendelig biokjemisk nettverk som er i stand til å generere, modulere, variere og spre seg, selv på avstand, millioner og millioner av informasjon.
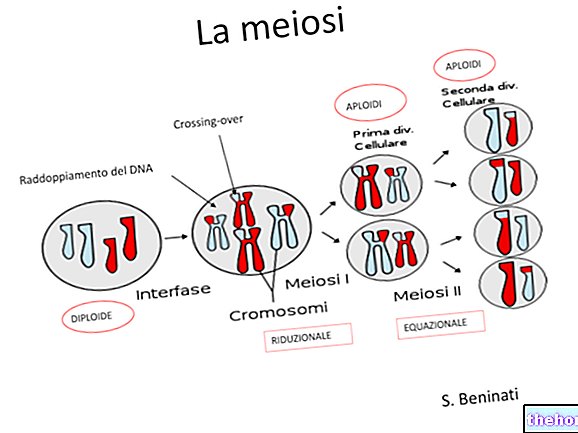
Hver celle i kroppen samhandler konstant med ECM, både under de mekaniske og kjemiske og energiske aspektene, med "dramatiske" effekter på vevets statiske og dynamiske arkitektur. Fibroblaster, for eksempel, virker vesentlig på kollagenet de produserer ved kontinuerlig behandling av det, for å kunne komprimere og forberede det i de former og mengder som kreves av det spesifikke organiske funksjonelle stedet. Hvis to små fragmenter av embryonalt vev er plassert langt fra hverandre, men i den samme kulturen av kollagengel, ser vi først en dannelse av perfekt justerte neokollagenfibre, som forbinder de to stubbene. Deretter migrerer fibroblastene ut av de to fragmentene langs neokollagenfibrene, kontrollerer avsetningen og blir overvåket etter tur. Dette funksjonelle syncytium er sannsynligvis tilstede under regenerative prosesser av ECM og utgjør et flerårig funksjonelt kontinuum som er i stand til å selvregulere ved fysiske forhold å klare de konstante variasjonene som kreves av de forskjellige vitale vevsfunksjonene (Albergati, 2004).
I følge P. A. Bacci representerer den interstitielle matrisen virkelig moren til vitale reaksjoner, stedet der først og fremst utvekslingen mellom materie og energi finner sted. Alle vev er forbundet og funksjonelt integrert med hverandre ikke i lukkede, men åpne systemer; kontinuerlige utvekslinger finner sted mellom dem, som kan skje både lokalt og systemisk, og utnytter biokjemiske, biofysiske og elektromagnetiske meldinger, det vil si ved bruk av de forskjellige energiformene. Den ioniske sammensetningen av det ekstracellulære interstitielle rommet utgjør et grunnleggende stoff som ikke bare tillater utveksling og liv, men også virker på genuttrykket til hver celle.
Som F. G. Albergati uttaler, representerer cellen og den ekstracellulære matrisen to verdener som tilsynelatende er atskilte, som nødvendigvis for hele levetiden, hvert øyeblikk, må samhandle for å fungere på en korrekt og synergistisk måte. Dette krever en ekstraordinær serie signaler etterfulgt av en like utrolig rekke molekylærbiologiske aktiviteter.
Ombygging av MEC
ECM må betraktes som en struktur i flerårig og konstant morfofunksjonell "ombygging", både under fysiologiske og patologiske forhold, basert på de funksjonelle kravene som kommer både fra sitt eget indre (gjennom "virkningen av metalloproteaser" og fra cellene ( for "virkningen av de mange proteinfraksjonene av vedheft). Redusert eller fraværende ECM -ombyggingskapasitet er dødelig for cellen. Som vi har sett, uttrykkes alle mobilfunksjoner i kraft av den tilsvarende funksjonen til strukturene til ECM, og de patologiske prosessene kan være primære eller sekundære til modifikasjonene av ECM.
Tydelige eksempler på ECM-ombygging er reparasjon av de dermal-hypodermiske lagene (denne prosessen krever presise sekvenser av matriksnedbrytning, migrering av spesifikke celler på dette stedet, syntese av en midlertidig matrise sammensatt av fibronektin, fibriner og store mengder type III-kollagen, fase med ombygging av den foreløpige matrisen som er grunnleggende for funksjonell restaurering av komponentene og følgelig for strukturell restaurering av selve vevet) og neoangiogenese (fysiologisk prosess for dannelse av nye kapillærer i vev og organer i forskjellige patologiske situasjoner, inkludert onkologisk). Neoangiogenese studeres for terapeutiske formål med det formål å gjenopprette korrekt blodtilførsel i iskemiske vev, for eksempel i hjertemuskelen eller i den perifere sirkulasjonen, samt å hemme prosessen, for eksempel i svulstfeltet (Shishido et al, 2003 )
De metalloprotease (MMP) er en familie av sink- og kalsiumholdige endopeptidaser som viser potensialet til å bryte ned alle proteiner og proteoglykankomponenter i ECM. De har sekvenser som ligner på interstitial kollagen og plasseres på ytre side av cellemembranen i en inaktiv (aktivert etter behov) form. Aktiviteten deres hemmes av et spesifikt middel kalt TIMP (Tissue Inhibitors of MetalloProteases).
Tidligere kalt kollagenase eller gelatinase for sin egenskap ved å denaturere naturlig eller denaturert kollagen (gelatin) - Birkedal -Hansen. Disse enzymene er involvert i mange fysiologiske og patologiske prosesser, inkludert angiogenese, embryogenese, inflammatoriske responser, åreforkalkning og mange artrheumatiske sykdommer (inkludert revmatoid artritt - Dieppe, 1995).
Andre artikler om "Viktigheten av den ekstracellulære matrisen i cellulær likevekt"
- Fibronektin, glukosaminoglykaner og proteoglykaner
- Ekstracellulær matrise
- Kollagen og elastin, kollagenfibre i den ekstracellulære matrisen
- Endringer av den ekstracellulære matrisen og patologier
- Bindevev og ekstracellulær matrise
- Deep fascia - Bindevev
- Fasciale mekanoreceptorer og myofibroblaster
- Deep fascia biomekanikk
- Holdning og dynamisk balanse
- Tensegrity og spiralformede bevegelser
- Nedre lemmer og kroppsbevegelse
- Setestøtte og stomatognatisk apparat
- Kliniske tilfeller, endringer i stillingen
- Kliniske tilfeller, holdning
- Postural evaluering - Klinisk case
- Bibliografi - Fra den ekstracellulære matrisen til holdningen. Er tilkoblingssystemet vår sanne Deus ex machina?