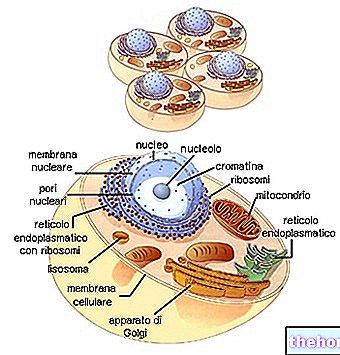
Mitose er konvensjonelt delt inn i fire perioder, kalt henholdsvis profase, metafase, anafase og telofase. De blir fulgt av oppdeling i to datterceller, kalt cytodierese.
Profas
I kjernen kan du gradvis se avgrensningen av fargbare filamenter, fremdeles forlenget og såret i en ball. Den gradvise spiraliseringen av DNA -strengene bundet til kjerneproteinene gjør dermed kromosomene gradvis identifiserbare. I mellomtiden forsvinner nukleolusen, mens sentriolen dobles. De to sentriolene vandrer til motsatte poler i kjernen, mens oppløsningen av kjernemembranen begynner. I øyeblikket av overgangen fra profase til metafase (siden noen identifiserer separat som prometafase) blir kromosomene forkortet og tydelig synlige, ikke lenger isolert i kjernemembranen; sentriolene er på motsatte poler, med en spindel av mikrotubuli som forbinder dem med meridianmote: kjernen har oppløst seg. Spindelen som forbinder sentriolene, kalt achromatisk spindel fordi den ikke kan farges (i motsetning til kromosomene), inkluderer både kontinuerlige fibre (spindelfibre), og fibre som i midtpunktet kobles til kromosomens sentromerer (kromosomfibre).

Metafase
I metafasen skiller spindelen seg tydelig ut, med alle kromosomene plassert på ekvatorialplanet, kalt ekvatorialplaten. I dette øyeblikket er kromosomene på maksimal forkortelse. Dette er øyeblikket der cellen er fikset for å telle og identifisere kromosomene. Hvert kromosom ser tydelig ut sammensatt av to identiske filamenter (kromatider), holdt sammen i et punkt som kalles sentromere ( de to kromatidene er et resultat av reduplisering). Sentromeren er det eneste kontaktpunktet, som en slags vedheft mellom to blodplater. Alle sentromerer er festet til det sentrale punktet til de kromosomale fibrene i den akromatiske spindelen (det er derfor kromosomene er i ekvatorial stilling).
Anafase
På slutten av metafasen bemerkes det at hver sentromer dobler seg, med hver halvdel som vandrer langs spindelen i retning av sin respektive pol. På dette tidspunktet kromatidene, dratt av sine respektive sentromerer, skiller seg klart i to grupper: hvert således kromatid som er atskilt, har så å si blitt myndig: fra nå av er det et kromosom bestemt for den respektive dattercellen.
Telofase
Kromosomene separert i to identiske klynger går tilbake til å despiralisere og rekonstituerer kjernen til to nye celler; den achromatiske smelten oppløses.
Cytodierese
Cytoplasma deler seg også ved gradvis struping og tilskriver de to dattercellene de respektive proporsjonene av både volum og celleorganeller. Spesielt må hver dattercelle motta minst en mitokondrion, ettersom den har mottatt en sentriole (dette er som nevnt strukturer med sin egen genetiske kontinuitet).
Det skal bemerkes at i grønnsaksriket, selv om de generelle egenskapene til mitose blir respektert, er det noen forskjeller. Først og fremst mangler sentriolene: ved spindelens poler er det optisk tomme mellomrom, kalt sentrosomer, som mikrotubuli utstråler. Videre på tidspunktet for cytodierese, der tildeling av et plastid (på grunn av dens genetiske kontinuitet) også må sikres til hver dattercelle, skjer separasjonen av dattercellene ikke ved kvelning, men ved dannelse av et septum, først bare av plasmalemma, deretter med påfølgende interposisjon av celleveggen.
Det genetiske grunnlaget representeres av "vekslingen mellom en dobling av det genetiske materialet (reduksjon av DNA, det vil si dobling av hvert kromosom til to like kromatider, forbundet gjennom sentromerer), og en halvering (separasjon av sentromerer, migrasjon av de to kromatidene i motsatt retning for å utgjøre de to nye like kjernene).
Siden, som vi vil se, er kromosomene tilstede i par av homologer (som kommer fra gameter), ser vi at antallet kromosomstrenger umiddelbart etter divisjonen er ett par for hver type kromosom. Ved å kalle antallet forskjellige kromosomtyper som er karakteristisk for de enkelte artene, er det normale kromosom -settet etter mitose n par homologe kromosomer (2n kromosomer = diploide celler).
Etter S -fasen vil imidlertid hvert kromosom ha doblet seg. Faktisk er det nødvendig å ha 4n kromatider for å kunne gi hver dattercelle 2n kromosomer. På denne måten ser vi at reduplisering og mitose veksler mellom 4n og 2n DNA -tråder.