Nukleinsyrer er kjemiske forbindelser av stor biologisk betydning; alle levende organismer inneholder nukleinsyrer i form av DNA og RNA (henholdsvis deoksyribonukleinsyre og ribonukleinsyre). Nukleinsyrer er svært viktige molekyler fordi de utøver primær kontroll over de grunnleggende livsprosessene i alle organismer.
Alt tyder på at nukleinsyrer har spilt en identisk rolle siden de første formene for primitivt liv som var i stand til å overleve (for eksempel bakterier).
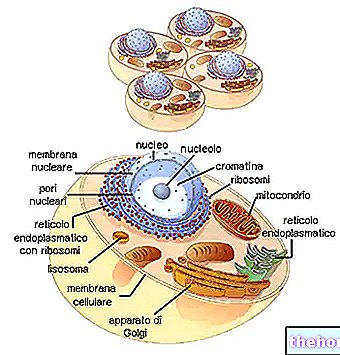
I cellene til levende organismer er DNA hovedsakelig tilstede i kromosomer (i delende celler) og i kromatin (i intercynetiske celler).
Den er også tilstede utenfor kjernen (spesielt i mitokondriene og plastidene, hvor den utfører sin funksjon som et informasjonssenter for syntese av deler av eller hele organellen).
RNA, derimot, er tilstede både i kjernen og i cytoplasma: i kjernen er den mer konsentrert i kjernen; i cytoplasma er den mer konsentrert i polysomene.
Den kjemiske strukturen til nukleinsyrer er ganske kompleks; de dannes av nukleotider, som hver (som vi har sett) dannes av tre komponenter: karbonhydrat (pentose), nitrogenbase (purin eller pyrimidin) og fosforsyre.
Nukleinsyrer er derfor lange polynukleotider, som skyldes sammenkoblingen av enheter som kalles nukleotider. Forskjellen mellom DNA og RNA ligger i pentosen og basen. Det er to typer pentose, en for hver type nukleinsyre:
1) Ribose i RNA;
2) Dessosiribose i DNA.
Når det gjelder basene, må vi gjenta skillet; pyrimidinbaser inkluderer:
1) cytosin;
2) Tymin, bare tilstede i DNA;
3) Uracil, kun tilstede i RNA.
Purinbaser, derimot, består av:
1) Adenin
2) Guanine.
Oppsummert finner vi i DNA: Cytosine - Adenine - Guanine - Thymine (C -A -G -T); mens vi i RNA har: Cytosine - Adenine - Guanine - Uracil (C -A -G -U).
Alle nukleinsyrer har polynukleotid lineær kjedestruktur; spesifisiteten til informasjonen er gitt av den forskjellige sekvensen av basene.
DNA -struktur
Nukleotidene i DNA -kjeden er knyttet til en esterbinding mellom fosforsyre og pentose; syren er bundet til karbon 3 i nukleotidpentosen og til karbon 5 i den neste; i disse bindingene bruker den to av de tre syregruppene; den resterende syregruppen gir molekylet sin sure karakter og lar den danne bindinger med basiske proteiner .
DNA har en dobbel spiralstruktur: to komplementære kjeder, hvorav den ene "går ned" og "den andre" går opp. "Til dette arrangementet tilsvarer begrepet" antiparallelle "kjeder, det vil si parallelle men med motsatte retninger. den ene siden begynner en av kjedene med en binding mellom fosforsyre og karbon 5 i pentosen og ender med et fritt karbon 3; mens retningen til den komplementære kjeden er motsatt. Vi ser også at hydrogenbindinger mellom disse to kjedene forekommer bare mellom en purinbase og en pyrimidin og omvendt, det vil si mellom Adenine og Thymine og mellom Cytosine og Guanine, og omvendt; det er to hydrogenbindinger i AT -paret, mens det i GC -paret er tre bindinger. Dette betyr at det andre paret har større stabilitet.
DNA -reduksjon
Som allerede nevnt med hensyn til den intercynetiske kjernen, kan DNA være i de "autosyntetiske" og "allosyntetiske" fasene, det vil si at de er engasjert i å syntetisere par av seg selv (autosyntese) eller et "annet stoff (RNA: allosyntese). Den er delt inn i tre faser, kalt G1, S, G2. I fase G1 (der G kan tas som den første veksten) syntetiserer cellen, under kontroll av kjernefysisk DNA, alt som trengs for metabolismen. I fase S (hvor S står for syntese, dvs. syntese av nytt kjernefysisk DNA) finner DNA -reduksjon sted. I fase G2 gjenopptar cellen veksten og forbereder seg på neste divisjon.
La oss kort se fenomenet som skjedde i fase S
Først og fremst kan vi representere de to antiparallelle kjedene som om de allerede var "despiraliserte". Fra en ekstrem brytes bindingene mellom basepar (A - T og G - C), og de to komplementære kjedene beveger seg fra hverandre (sammenligningen av åpningen til en "blits" er egnet). På dette tidspunktet er et enzym ( DNA-polymerase) "flyter" langs hver enkelt kjede, noe som favoriserer dannelsen av bindinger mellom nukleotidene som komponerer det og nye nukleotider (tidligere "aktivert" med energi frigjort av "ATP) som er utbredt i karyoplasma. En ny timina er nødvendigvis knyttet til hver adenin, og så videre, og danner gradvis en ny dobbeltkjede fra hver enkelt kjede.
DNA-polymerasen ser ut til å virke in vivo likegyldig på de to kjedene, uansett "retning" (fra 3 til 5 eller omvendt). På denne måten, når hele den opprinnelige doble DNA-kjeden er dekket, to dobbeltkjeder, nøyaktig det samme som originalen. Begrepet som definerer dette fenomenet er "semikonservativ reduplisering", hvor "reduplisering" konsentrerer betydningen av kvantitativ dobling og eksakt kopi, mens "semikonservativ" minner om det faktum at for hver ny dobbeltkjede av DNA bare en kjede er nyintetisk.
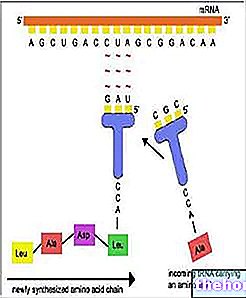
DNA inneholder genetisk informasjon, som den overfører til RNA; sistnevnte i sin tur overfører den til proteiner, og regulerer dermed cellens metabolske funksjoner. Følgelig er hele metabolismen direkte eller indirekte under kontroll av kjernen.
Den genetiske arven vi finner i DNA er bestemt til å gi spesifikke proteiner til cellen.
Hvis vi tar dem i par, vil de fire basene gi 16 mulige kombinasjoner, det vil si 16 bokstaver, ikke nok for alle aminosyrer. Hvis vi i stedet tar dem i trillinger, vil det være 64 kombinasjoner, som kan virke for mange, men som i virkeligheten alle er i bruk ettersom vitenskapen har oppdaget at forskjellige aminosyrer er kodet av mer enn en trilling. Vi har derfor oversettelsen fra de 4 bokstavene i de nitrogenholdige basene til nukleotidene til 21 av aminosyrene; Imidlertid, før "oversettelsen", er c "" transkripsjonen ", fremdeles innenfor" fire bokstaver "-konteksten, det vil si passering av den genetiske informasjonen fra de fire bokstavene i DNA til de fire bokstavene i RNA, idet ta hensyn til at c, i stedet for det sjenerte (DNA), er "uracil" (RNA).
Transkripsjonsprosessen skjer når, i nærvær av ribonukleotider, enzymer (RNA-polymerase) og energi som finnes i ATP-molekylene, åpnes DNA-kjeden og RNA syntetiseres, som er en trofast gjengivelse av den genetiske informasjonen. åpen kjede.
Det er tre hovedtyper av RNA, og de stammer alle fra kjernefysisk DNA:
- RNAm (messenger)
- RNAr (ribosomal)
- RNAt eller RNA (overføring eller løselig)