For å snakke om de tjue aminosyrene som består av proteinstrukturene og de modifiserte, ville det være nødvendig å beskrive minst tolv spesialiserte metabolske veier.
Men hvorfor bruker celler så mange metabolske veier som krever energi (for eksempel for å regenerere de katalytiske stedene til enzymer), hver med et enzymatisk arv, for å katabolisere aminosyrer? Fra nesten alle aminosyrer er det mulig å oppnå, via spesialiserte veier, metabolitter som i liten grad brukes til å produsere energi (for eksempel gjennom glukoneogenese og banen til ketonlegemer), men som fremfor alt fører til dannelse av kompleks molekyler, med et stort antall karbonatomer (for eksempel fra fenylalanin og tyrosin, produseres hormoner i binyrene som er spesialiserte for dette formålet); hvis det på den ene siden ville være enkelt å produsere energi fra aminosyrer, på den andre siden ville det være komplisert å bygge komplekse molekyler med utgangspunkt i små molekyler: katabolismen av aminosyrer gjør at de kan utnytte skjelettet sitt for å skaffe større arter.
To eller tre hektar aminosyrer nedbrytes daglig av et sunt individ: 60-100 g av dem stammer fra proteinene som ble introdusert med dietten, men mer enn 2 hektar hentes fra normal omsetning av proteiner som er en integrert del av organismen ( aminosyrer av disse proteinene, som er skadet av redokseprosesser, erstattes av andre og kataboliseres).
Aminosyrer gir et energibidrag når det gjelder ATP: Etter å ha fjernet a-aminogruppen, kan det gjenværende karbonholdige skjelettet av aminosyrer, etter passende transformasjoner, gå inn i krebs-syklusen. Når tilførsel av næringsstoffer mangler og mengden glukose reduseres, aktiveres glukoneogenese: glukoneogenetiske aminosyrer sies å være de som etter passende modifikasjoner kan innføres i glukoneogenese; glukoneogenetiske aminosyrer er de som kan omdannes til pyruvat eller i fumarat (fumarat kan omdannes til malat som forlater mitokondrion og i cytoplasma omdannes til oxaloacetat hvorfra fosfoenolpyruvat kan hentes) eddik-acetat.
Den som nettopp er beskrevet er et veldig viktig aspekt fordi aminosyrer kan rette opp en sukkermangel ved umiddelbar faste; hvis fasten vedvarer, etter to dager griper lipidmetabolismen inn (fordi proteinstrukturene ikke kan angripes for mye), er det i denne fasen at siden glukoneogenesen er svært begrenset, blir fettsyrene omdannet til acetylkoenzym A og ketonlegemer. Fra ytterligere faste tilpasser hjernen seg også til å bruke ketonlegemene.
Overføringen av a-aminogruppen fra aminosyrer skjer gjennom en transamineringsreaksjon; enzymene som katalyserer denne reaksjonen, de sier faktisk transaminaser (eller aminotransferase). Disse enzymene bruker en enzymatisk kofaktor kalt pyridoksalfosfat, som griper inn i aldehydgruppen. Pyridoksalfosfat er produktet av fosforyleringen av pyridoksin, som er et vitamin (B6) som hovedsakelig finnes i grønnsaker.
Transaminaser har følgende egenskaper:
Høy spesifisitet for et ketoglutarat-glutamat α-par;
De er oppkalt etter det andre paret.
Transaminaseenzymer involverer alltid a-ketoglutarat-glutamatparet og skilles ut fra det andre paret som er involvert.
Eksempler:
L "aspartat transaminase dvs. GOT (Glutamat-Ossalacetat-transaminase): enzymet overfører α-aminogruppen fra aspartat til α-ketoglutarat, og får oxaloacetat og glutamat.
L "alanintransaminase dvs. GTP (Glutamat-Pyruvat-transaminase): enzymet overfører a-aminogruppen fra "alanin til" α-ketoglutarat, og får pyruvat og glutamat.
De forskjellige transaminasene bruker α-ketoglurat som en acceptor for aminogruppen av aminosyrer og omdanner det til glutamat; mens aminosyrene som dannes brukes i banen til ketonlegemene.
Denne reaksjonstypen kan skje i begge retninger siden de bryter og danner bindinger med samme energiinnhold.
Transaminasene er både i cytoplasma og i mitokondrion (de er stort sett aktive i cytoplasma) og er forskjellige i sitt isoelektriske punkt.
Transaminasene er også i stand til å dekarboksylere aminosyrer.
Det må være en måte å konvertere glutamat tilbake til α-ketoglutarat: dette gjøres ved deaminering.
Der glutamat dehydrogenase det er et enzym som er i stand til å transformere glutamat til α-ketoglutarat og derfor konvertere aminogruppene til aminosyrer som finnes i form av glutamat til ammoniakk. Det som skjer er en redokseprosess som passerer gjennom det mellomliggende a-aminoglutarat: ammoniakk og α-ketoglutarat frigjøres og går tilbake til sirkulasjonen.
Avhendingen av aminogruppene til aminosyrene passerer deretter transaminasene (som varierer etter substratet) og glutamatdehydrogenasen, som bestemmer dannelsen av ammoniakk.
Det er to typer glutamatdehydrogenase: cytoplasmatisk og mitokondrielt; kofaktoren, som også er cosubstratet til dette enzymet er NAD (P) +: glutamat dehydrogenase bruker enten NAD + eller NADP + som en akseptor for reduserende effekt. Den cytoplasmiske formen foretrekker, men ikke utelukkende, NADP + mens den mitokondrielle formen foretrekker NAD +. Den mitokondrielle formen har til hensikt å avhende aminogrupper: den fører til dannelse av ammoniakk (som er et substrat for et spesialisert enzym i mitokondrien) og NADH (som sendes til respirasjonskjeden). Den cytoplasmatiske formen virker i motsatt retning, det vil si at den bruker ammoniakk og α-ketoglutarat for å gi glutamat (som har en biosyntetisk destinasjon): denne reaksjonen er en reduktiv biosyntese og kofaktoren som brukes er NADPH.
Glutamat dehydrogenase virker når det er nødvendig å kvitte seg med aminogruppene i aminosyrer som ammoniakk (via urin) eller når skjelettene til aminosyrer er nødvendige for å produsere energi: dette enzymet vil derfor ha negative modulatorer systemene som er en indikasjon med god energitilgjengelighet (ATP, GTP og NAD (P) H) og som positive modulatorer, systemene som indikerer behov for energi (AMP, ADP, BNP, NAD (P) +, aminosyrer og skjoldbruskhormoner).
Aminosyrer (hovedsakelig leucin) er positive modulatorer av glutamatdehydrogenase: hvis aminosyrer er tilstede i cytoplasma, kan de brukes til proteinsyntese, eller de må kastes fordi de ikke kan akkumuleres (dette forklarer hvorfor aminosyrer er positive modulatorer ).
Deponering av ammoniakk: ureasyklus
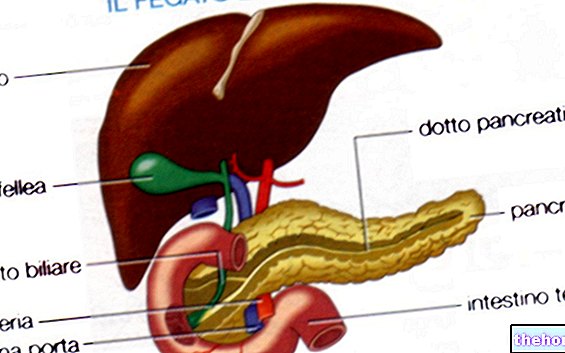
Fisk avhender ammoniakk ved å føre det inn i vann gjennom gjellene; fugler omdanner det til urinsyre (som er et kondensasjonsprodukt) og eliminerer det med avføring. La oss se hva som skjer hos mennesker: vi har sagt at glutamat dehydrogenase omdanner glutamat i α- ketoglutarat og ammoniakk, men vi har ikke sagt at dette bare skjer i mitokondriene i leveren.
En grunnleggende rolle for avhending av ammoniakk gjennom ureasyklusen, spilles av mitokondrielle transaminaser.

Karbondioksid, i form av bikarbonation (HCO3-), aktiveres av biotinkofaktoren som danner karboksybiotin som reagerer med ammoniakk for å gi karbaminsyre; den neste reaksjonen bruker ATP for å overføre et fosfat til karbaminsyren som danner karbamylfosfat og ADP (omdannelsen av ATP til ADP er drivkraften for å få karboksybiotin.) Denne fasen katalyseres av karbamylfosfatsyntetase og forekommer i mitokondrien. Karbamylfosfat og ornitin er substrater for enzymet ornitintrans karbamylase som konverterer dem til citrullin; denne reaksjonen skjer i mitokondriene (hepatocytter). Den produserte sitrullinen forlater mitokondrien og går i cytoplasma under "virkningen av"argininsuksinatsyntetase: det er sammensmeltningen mellom det karbonholdige skjelettet til citrullin og et aspartat gjennom et nukleofilt angrep og påfølgende eliminering av vann. Argininsuccinatsyntetaseenzymet krever et ATP -molekyl, så det er en energisk kobling: hydrolysen av ATP til AMP og pyrofosfat (sistnevnte omdannes deretter til to molekyler ortofosfat) skjer ved utvisning av et molekyl d "vann fra substratet og ikke ved virkningen av vannet i mediet.
Det "neste enzymet er"argininsuccinase: Dette enzymet er i stand til å dele argininsuksinat til arginin og fumarat i cytoplasma.
Ureasyklusen fullføres av enzymet arginase: urea og ornitin oppnås; urea kasseres av nyrene (urin) mens ornitin går tilbake til mitokondrien og gjenopptar syklusen.
Urea -syklusen er gjenstand for indirekte modulering av arginin: akkumulering av arginin indikerer at ureasyklusen må fremskyndes; moduleringen av arginin er indirekte fordi arginin positivt modulerer acetylglutamatsyntetase -enzymet. Sistnevnte er i stand til å overføre en acetylgruppe på nitrogenet til et glutamat: N-acetylglutamat dannes som er en direkte modulator av karbamyl-fosfosyntetaseenzymet.
Arginin akkumuleres som en metabolitt i ureasyklusen hvis produksjonen av karbamylfosfat ikke er tilstrekkelig til å kvitte seg med ornitinet.
Urea produseres bare i leveren, men det er andre steder der de første reaksjonene finner sted.
Hjernen og musklene bruker spesielle strategier for å eliminere aminogrupper. Hjernen bruker en veldig effektiv metode der et enzym brukes glutaminsyntetase og et enzym glutamase: den første finnes i nevroner, mens den andre finnes i leveren. Denne mekanismen er veldig effektiv av to grunner:
To aminogrupper transporteres fra hjernen til leveren i et enkelt kjøretøy;
Glutamin er mye mindre giftig enn glutamat (glutamat utfører også neuronal overføring og må ikke overstige den fysiologiske konsentrasjonen).
Hos fisk bringer en lignende mekanisme aminogruppen av aminosyrer til gjellene.
Fra muskelen (skjelett og hjerte) når aminogruppene leveren gjennom glukose-alaninsyklusen; enzymet som er involvert er glutamin-pyruvat-transaminase: det tillater transponering av aminogrupper (som er i form av glutamat), omdanner pyruvat til alanin og samtidig glutamat til α-ketoglutarat i muskelen og katalyserer omvendt prosess i leveren.
Transaminaser med forskjellige oppgaver eller posisjoner har også strukturelle forskjeller og kan bestemmes ved elektroforese (de har forskjellige isoelektriske punkter).
Tilstedeværelsen av transaminaser i blodet kan være et symptom på lever- eller hjerteskade (dvs. vevsskade på lever eller hjerteceller); transaminasene er i svært høye konsentrasjoner både i leveren og i hjertet: gjennom elektroforese er det mulig å fastslå om skaden har oppstått i leveren eller hjertecellene.








-cos-cause-e-disturbi-associati.jpg)