Tidligere ble det ortosympatiske systemet kalt "ergo tropico"; dens aktivering bestemmer faktisk sløsing med energi som lett blir gjort tilgjengelig ved nedbrytning av glykogen til glukose, ved hydrolyse av lipider og ved akselerasjon av hjerteaktivitet; på denne måten forbereder organismen seg på å reagere på en tilstand av alvorlig stress , traumer, plutselige temperaturendringer eller alvorlig fysisk anstrengelse ("kamp eller fluktreaksjon"). Denne umiddelbare reaksjonen på en ugunstig tilstand er mulig fordi den sympatiske personen generelt utfører sin handling på en utbredt måte.
Det parasympatiske systemet ble kalt "trofotropisk" fordi det, i motsetning til det ortosympatiske, aktiveres under betingelser for gjenoppretting eller hvile og fordøyelse av organismen; Derfor spiller dette systemet en grunnleggende rolle for fordøyelsesfunksjoner, for utvinning av energireserver og for gjenoppretting av fysiologisk trykk og hjertesykdommer. Responsen som følge av aktiveringen av det parasympatiske kalles "sektoriell type", det vil si at det påvirker et "lokalisert område" av organismen. Det parasympatiske, med sin trofotropiske aktivitet, er derfor ansvarlig for å opprettholde organismens vitale funksjoner.
Under fysiologiske forhold er de orto- og parasympatiske funksjonene i likevekt, og eventuelle situasjoner med liten ubalanse blir fysiologisk korrigert gjennom "høyrefleksmekanismer", som - avhengig av tilfellet - tar sikte på henholdsvis å øke eller redusere "ortohandling. Og parasympatisk.

Overføringen av impulsen i de efferente veiene formidles av KOLINERGISKE pre-ganglioniske nevroner, uavhengig av om de er ortho eller parasympatiske: det vil si at de frigjør nevrotransmitteren Acetylcholine (Ach) på synaptisk nivå. Ach interagerer med de nikotiniske kanalreseptorene som er tilstede på ganglia; reseptorene som aktiveres, sender impulsen til post-ganglioniske fibre, som når effektororganet og frigjør: de som tilhører den parasympatiske nevrotransmitteren acetylkolin og de som tilhører det ortosympatiske Noradrenalin (Nor ).
Den somatiske innervasjonen, som styrer alle skjelettmuskulaturen, har nevronfibre uten ganglier, som stammer fra ryggmargen (motoriske nevroner i ryggraden), men også kolinerge; sistnevnte samhandler med "muskel" nikotinreseptorer, såkalt fordi de er lokalisert på skjelettmuskler Muskelnikotinreseptorer er forskjellige fra nikotinreseptorer som er tilstede på ganglier, derfor må legemidler som virker på disse reseptorene ha en selektiv virkning, ellers ville det være en risiko for å kompromittere hele den pre-ganglioniske sympatiske overføringen. Det må tas en egen diskusjon for binyremedullaen, hvis sympatiske innervering skiller seg fra alle andre organer fordi den mangler det post-ganglioniske nevronet; med andre ord, den pre-ganglioniske nevron-ganglionfrigjøringen Ach direkte på den nikotinreseptoren som er tilstede i adrenal medulla, som vil frigjøre nevrotransmitteren Adrenalin direkte inn i blodstrømmen, gjennom hvilken den når sine aktive steder ved å samhandle med de adrenerge reseptorene.
Turn-over av nevrotransmitterne i de orto- og parasympatiske systemene

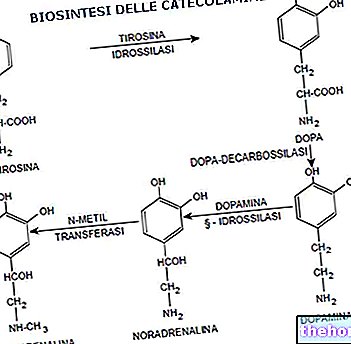
KATEKOLAMINER (adrenalin, noradrenalin og dopamin): syntetisert i de ortosympatiske post-ganglioniske nerveender ved transformasjon av aminosyren tyrosin til dopa av tyrosin-hydroksylase-enzymet, og deretter til dopamin av dopa-dekarboksylase-enzymet; dopamin lagres i synaptiske vesikler og til slutt omdannes til noradrenalin.

Dopamin i seg selv kan fungere som en nevrotransmitter, i så fall snakker vi om dopaminerge nevroner, som fremfor alt ligger på nivået av CNS. Vesiklene som inneholder nevrotransmitteren migrerer til cellemembranen etter depolarisering og frigjør noradrenalin på synaptisk nivå, hvor den interagerer med de respektive reseptorene. Etter å ha utført sin funksjon, blir noradrenalin tatt opp av nerveendene og nedbrytes av spesifikke enzymer, kalt mono-amino-oksidase eller MAO. I minimal grad, på synaptisk nivå, kan noradrenalin gjennomgå virkningen av COT (katecholamin transferase).
Andre artikler om "Neurotransmittere i det parasympatiske og ortosympatiske nervesystemet"
- Parasympatiske og ortosympatiske nervesystem
- Kolinomimetiske legemidler







.jpg)





---pan-di-spagna-per-rotolo-dolce.jpg)