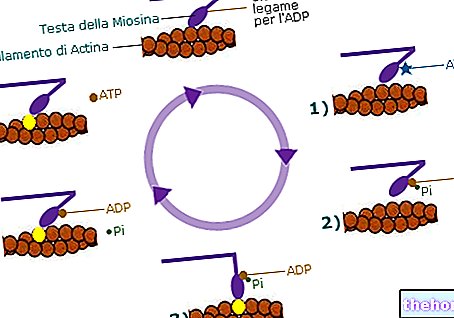
Krebs -syklusen kalles også trikarboksylsyresyklusen og bruker acetylkoezim A som startmetabolitt, som oppnås ved virkningen av pyruvatdehydrogenase på pyruvat som produseres ved glykolyse.
ATP og reduserende effekt oppnås fra krebs -syklusen; reduksjonskraften sendes til respirasjonskjeden hvor NADH og FADH2 oksideres henholdsvis til NAD + og FAD: reduksjonskraften overføres langs respirasjonskjeden til koblingssystemer som ytterligere ATP produseres fra.
Krebs -syklusen er et nervesenter ikke bare for glukosemetabolismen, men også for metabolismen av fettsyrer og aminosyrer, faktisk kommer pyruvat som omdannes til acetylkoenzym A ikke bare fra nedbrytning av glukose: det oppnås, for eksempel, også fra transaminering av alanin (en aminosyre).
Omtrent 80% av acetylkoenzym A som deltar i krebs -syklusen kommer fra metabolismen av fettsyrer.
Acetylkoenzym A er en tioester, derfor har den et høyt energiinnhold som utnyttes av sitratsyntase for å danne en ny karbon-karbonbinding; sitratsyntase er det første enzymet i krebs -syklusen.
Metylkarbonet av acetylkoenzym A frigjør villig (ved tautomerisme) et proton (blir til et karboanion) og angriper karbonylkarbonet av oksaloacetat: et tioester (citril -koenzym A) med høyt energiinnhold dannes, hvorfra sitratet oppnås ved hydrolyse. og koenzym A. reformeres. Sitratsyntasen moduleres negativt av produktet, det vil si av sitratet og av ATP: hvis sitratet akkumuleres betyr det at dette stadiet er raskere enn de andre, derfor må det bremse ned (sitrat er et negativt modulator).

ATP påvirker også virkningen av sitratsyntase siden reduksjonskraften oppnås fra krebs -syklusen som deretter sendes til respirasjonskjeden som ATP produseres fra; hvis ATP akkumuleres betyr det at det produseres mer enn det som trengs. Ved å bremse krebs-syklusen (syklusen bremses hvis et av stadiene blir bremset) reduseres også produksjonen av ATP: den negative moduleringen av ATP er en tilbakemeldingsmodulasjon (dannelsen av et av sluttproduktene er modulert ved å justere hastigheten til ett trinn i prosessen).
I den andre fasen av krebs -syklusen blir sitratet omdannet til isocitrat ved virkningen av enzymet akonitase; navnet på enzymet stammer fra det faktum at sitratet først dehydreres med dannelsen av cis-akonitat, og deretter kommer vannet inn igjen ved å feste seg til et karbon som er forskjellig fra det som det tidligere var bundet til. Isocitratet oppnås uten at substratet forlater det katalytiske stedet; akonitase er et stereospesifikt enzym: det gjenkjenner de tre karboksylsentrene til sitratet, og dette får sitratet til å forbli bundet til enzymet slik at utgangen og inntreden av "vannet alltid passerer gjennom cis-akonitat-mellomproduktet.
I den tredje fasen av krebs -syklusen er det den første energihensynet fordi det er tap av et karbon som elimineres som karbondioksid. Enzymet som katalyserer dette stadiet er isocitrat dehydrogenase; substratet gjennomgår først og fremst en dehydrogenering: NAD + får redusert effekt og oksalosuccinat dannes (det er et oksalderivat av ravsyre) .Oksalosuccinatet gjennomgår deretter dekarboksylering til α-ketoglutarat.
Enzymet isocitrat dehydrogenase har to modulasjonssteder: en positiv modulasjon på grunn av ADP og en negativ modulasjon på grunn av ATP. Mengden ATP som forbrukes daglig er veldig høy: ATP gir energien som frigjøres ved hydrolysen, "ADP og alt" ortofosfat.
Den totale konsentrasjonen av nukleosider (nitrogenbasert pluss sukker) og nukleotider (nuklosid pluss fosfat) i en organisme er nesten konstant: å si derfor at c "er mye ATP eller lite ADP (eller omvendt, mye ADP og lite ATP) er det samme; ADP er et synonym for behov for energi og er derfor en positiv modulator, mens ATP er et symptom på energitilgjengelighet og derfor er en negativ modulator.
FORTSETT: Andre del "