Redigert av Dr. Giovanni Chetta
ECM beskrives generelt som sammensatt av flere store klasser av biomolekyler:
- Strukturelle proteiner (kollagen og elastin)
- Spesialiserte proteiner (fibrillin, fibronektin, laminin etc.)
- Proteoglykaner (aggrecans, syndecans) og glusaminoglycans (hyaluronaner, kondroitinsulfater, heparansulfater, etc.)
Strukturelle proteiner
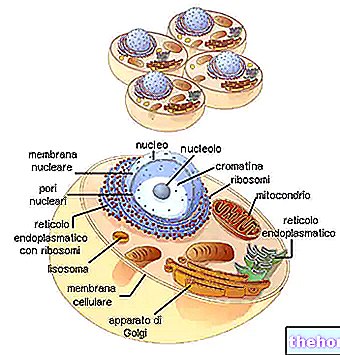
Kollagener danner den mest representerte familien av glykoproteiner i dyreriket. De er de mest tilstedeværende proteiner i den ekstracellulære matrisen (men ikke den viktigste) og er de grunnleggende bestanddelene i riktig bindevev (brusk, bein, fascia, sener, ledbånd).
Det er minst 16 forskjellige typer kollagen, hvorav type I, II og III er de mest tilstede på nivå med de typiske fibrillene (type IV danner en slags retikulum som representerer hovedkomponenten i basallamina).
Kollagener blir for det meste syntetisert av fibroblaster, men epitelceller er også i stand til å syntetisere dem.
Kollagenfibre samhandler kontinuerlig med en enorm mengde andre molekyler i den ekstracellulære matrisen, som utgjør et biologisk kontinuum som er grunnleggende for cellens liv.De tilhørende kollagenene i fibriller spiller en dominerende rolle i dannelsen og vedlikeholdet av strukturer som er i stand til å motstå spenningskrefter, er nesten uelastisk (glukosaminglykaner utfører en handling av motstand mot kompresjon). På en eller annen måte produseres og metaboliseres kollagen på nytt som en funksjon av den mekaniske belastningen og dens viskoelastiske egenskaper, som vi vil se i avsnittet "Viskoelastisitet av fascia ", en stor innvirkning på menneskets holdning. Som en ytterligere demonstrasjon av kollagens evne til å endre seg i henhold til miljøpåvirkning, forutsatt f.eks. varierende grader av stivhet, elastisitet og motstand, er det kollagener, definert med begrepet FACIT (Fibril Associated Collagen with Interrupted Triple helices) i stand til å fungere funksjonelt som proteoglykaner (beskrevet i avsnittet "Glukosaminoglykaner og proteoglykaner").
Kollagenfibrene, takket være belegg av PG / GAG (proteoglykaner / glukosaminoglykaner), har egenskaper til biosensorer og bioledere: de relative elektriske ladningene resulterer i en større evne til å binde vann og utveksle ioner, derfor en større elektrisk kapasitet.
Vi vet at enhver mekanisk kraft som er i stand til å generere en strukturell deformasjon, stresser de intermolekylære bindingene og gir en liten elektrisk strøm, det vil si piezoelektrisk strøm (Athenstaedt, 1969). I slike tilfeller fordeler kollagenfibrene de positive ladningene på deres konvekse overflate og de negative på den konkave overflaten, og transformerer dermed til halvledere (de tillater strøm av elektroner på enveisoverflaten). Siden den piezoelektriske energien (så vel som den pyroelektriske energien generert av termiske påkjenninger) blir nøytralisert av ionene i sirkulasjon på veldig kort tid (ca. 10-7-10-9 sekunder), vil arrangementet av PG / GAG på signalet er avgjørende for spredning av signaloverflaten til fibrillene, for eksempel å fungere som "repeatere" av den elektriske impulsen. Spesielt er en langsgående periodisitet på ca. 64 nm (som under det optiske mikroskopet vises som en strek) tillater en forplantningshastighet av impulsen lik ca. 64 m / s (tilsvarer ledningshastigheten til raske nervefibre) - Rengling, 2001. Det sterke dipolare øyeblikket av kollagenfibriller og deres resonanskapasitet (egenskap som er felles for alle peptidstrukturer), så vel som den lave dielektriske konstanten til MEC, letter overføringen av elektromagnetiske signaler.Derfor besitter det tredimensjonale og allestedsnærværende kollagennettverket også den særegne egenskapen ved å lede bioelektriske signaler i de tre dimensjonene til rommet, basert på det relative arrangementet mellom kollagenfibriller og celler, i afferent retning (fra ECM til cellene) eller, omvendt, efferent.
Alt dette representerer et sanntids MEC-celle kommunikasjonssystem, og slike elektromagnetiske bio-signaler kan føre til viktige biokjemiske endringer, for eksempel i bein kan ikke osteoklaster "fordøye" piezoelektrisk ladet bein (Oschman, 2000).
Til slutt bør det understrekes at cellen, ikke overraskende, produserer kontinuerlig og med et betydelig energiforbruk (ca. 70%) materiale som nødvendigvis må utvises, hovedsakelig gjennom eksklusiv lagring av protokollagen (biologisk forløper til kollagen) i spesifikke vesikler (Albergati, 2004).
De aller fleste virveldyrvev krever samtidig tilstedeværelse av to vitale egenskaper: styrke og elastisitet. Et ekte nettverk av elastiske fibre, plassert inne i ECM i disse vevene, gjør det mulig å gå tilbake til de første forholdene etter sterke trekk. De elastiske fibrene er i stand til å øke forlengelsen av et organ eller en del av det med minst fem ganger. Lange, uelastiske kollagenfibre er spredt mellom de elastiske fibrene med den presise oppgaven å begrense "overdreven deformasjon på grunn av trekk i vevene. L"elastin representerer hovedkomponenten i elastiske fibre. Det er et ekstremt hydrofobt protein, omtrent 750 aminosyrer i lengde, ettersom kollagen er rikt på prolin og glycin, men i motsetning til kollagen er det ikke glykert og inneholder mange hydroksyprolinrester og ikke hydroksylisin. Elastin fremstår som et ekte biokjemisk nettverk av uregelmessig tredimensjonal form, sammensatt av fibre og lameller som gjennomsyrer ECM for alle bindevev. Det finnes i spesielt store mengder i blodårene med elastiske egenskaper (det er proteinet av ECM mer tilstede i arteriene og representerer mer enn 50% av den totale tørrvekten til aorta), i leddbåndene, i lungen og i huden. I dermis, i motsetning til det som skjer med kollagen, har tetthet og volum av elastin en tendens til å øke over tid, men det gamle elastinet virker generelt hovent, nesten hovent, ofte med et fragmentert utseende og med en reduksjon i komponenten. "Amorf" (Pasquali Rochetti et al, 2004). Glatte muskelceller og fibroblaster er de viktigste produsentene av forløperen, tropoelastin, som skilles ut i de ekstracellulære mellomrommene.
Andre artikler om "Kollagen og elastin, kollagenfibre i den ekstracellulære matrisen"
- Ekstracellulær matrise
- Fibronektin, glukosaminoglykaner og proteoglykaner
- Betydningen av den ekstracellulære matrisen i cellulær likevekt
- Endringer av den ekstracellulære matrisen og patologier
- Bindevev og ekstracellulær matrise
- Deep fascia - Bindevev
- Fasciale mekanoreceptorer og myofibroblaster
- Deep fascia biomekanikk
- Holdning og dynamisk balanse
- Tensegrity og spiralformede bevegelser
- Nedre lemmer og kroppsbevegelse
- Setestøtte og stomatognatisk apparat
- Kliniske tilfeller, endringer i stillingen
- Kliniske tilfeller, holdning
- Postural evaluering - Klinisk case
- Bibliografi - Fra den ekstracellulære matrisen til holdningen. Er tilkoblingssystemet vår sanne Deus ex machina?













-cos-cause-e-rimedi.jpg)